![]()
![]()
![]()
The shell of Pinctada fucata is about 1.5 mm thick over the greater part. The growth edge or projecting lamellae are laid down by the oyster at successive intervals on the distal border. The non-nacreous border of the inner surface of the valves are characterized by brownish or reddish patches which coincide with the external rays.
The adductor scar is elongated and sub-central. The pallial line and scars are caused by the insertion of the pallial muscles in fan-shaped bundles of fibres radiating outwards. There are 12–15 insertion scars between the umbo and antero-ventral border. Besides these distinct scars, there is a narrow continuous insertion band confluent with the posterior and ventral edges of the adductor scars. Its scar merges with that of the adductor scar. The hinge is narrow and runs along the greater part of the straight dorsal edge. Elongated ridge-like teeth are present at the anterior and posterior ends of the ligament.
The shell is composed of three layers. The very thin outer layer is uncalcified cuticular conchiolin layer or periostracum. This is an extremely delicate horny layer which allows the colour of the layer below to show through and usually becomes worn off in old shells. At the free margin of the shell the periostracum is very thin, transparent and extends beyond the calcareous matter. It is reflected to join the surface of the ectoderm cells of the mantle edge in the longitudinal groove where it is secreted.
The middle or prismatic layer shows a cellular structure formed of calcareous prisms or columns running vertically to the surface and appearing polygonal in section. The prismatic layer is deposited by the mantle epithelium near the free edge just behind the margin which forms the periostracum and many such layers on fusion are formed successively, each new one beneath the last, as the shell grows. The innermost nacreous or mother-of-pearl layer is composed of numerous fine lamellae of aragonite crystals (Fig. 2).
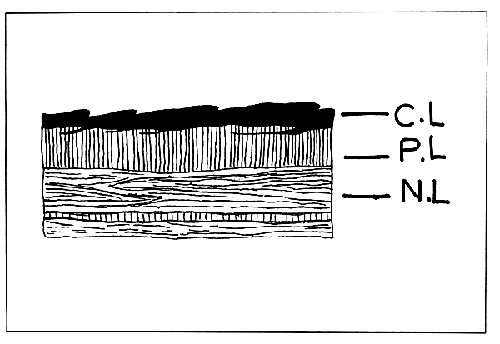
FIGURE 2. Section of the shell of Pinctada fucata. C.L.= conchiolin layer; P.L.= prismatic layer; and N.L.= nacreous layer.
The free edge of the mantle lobe is thick, pigmented and fringed with branched tentacles. The pallial edge of the mantle is attached to the shell, a little away from the margin. Each pallial lobe may be divided into three parts, the central, distal or muscular and marginal mantle.
The foot is highly mobile, tongue-shaped organ capable of considerable elongation and contraction (Fig. 3). It arises from the anterior region of the visceral mass nearly midway between the mouth and the intestinal lobe and the anterior branchiae flanking it on either side. The dorsal portion has more chromatopores while the ventral portion has the pedal groove. The major part of the foot is composed of a network of fibres running in various directions, thus ensuring a wide range of movement. It is extensively penetrated by blood spaces and the organ is highly contractible.
The byssus gland organ (Fig. 3) is located ventrally at the proximal end of the foot. The byssal gland lodges the common root of a bundle of stout, laterally compressed bronze-green fibres: the byssal threads. Each fibre of the byssus anchors the pearl oyster to rocks and other hard objects by means of a discoid attachment at the distal extremity. The anterior edge of the mouth of the byssal gland passes into the pedal groove (Fig. 3) extending along the whole of the remaining length of the ventral surface of the foot.
The pearl oyster is monomyarian, possessing only the
posterior adductor (Fig. 3), the largest and the most
important muscle in its body. The adductor muscle
stretches transversely across the body from valve to valve.
It is a massive wedge-shaped bundle of muscle fibres. The
narrow end points upwards and lies immediately behind the
ventricle of the heart. The terminal part of the rectum
runs in the middle line along the posterior surface. It
has two distinct regions, one a narrow tendonous strip made
up of white glistening fibres forming the posterior border,
the other composed of broad and massive semitranslucent
fibres occupying the remainder of the mass. The power
exerted by the adductor in bringing the valves together by
its contraction is considerable with rapid action
resembling a ratchet mechanism.
The retractors of the foot are a pair of symmetrically
disposed muscles lying in the horizontal plane of the body.
The muscles are V-shaped and originate from the byssal gland. Their ends are attached to the right and left valves without making a separate scar in the nacre.
The foot has four levators, two anterior and two posterior. The contraction of the anterior levator causes the foot to be retracted and raised dorsally. The posterior levators are short and insignificant, originating at level with the mouth, passing through the visceral mass to be attached to the valves behind the anterior levator scar. The branchial muscles cause the shortening of the gills and withdrawal of their posterior extremities. They run within each ctenidial axis from end to end, close to the dorsal edge. There are also muscle bundles running longitudinally down on each side of the principal filaments. The pallial muscles are retractable and together they constitute the orbicular muscle of the mantle. With the exception of heart and indistinct striations on larger portion of the adductor muscle, the muscle fibres throughout the oyster's body are non-striped.
The oesophagus, stomach, and the greater portion of the intestine lie within the viscero-pedal mass. Two horizontal lips, the labial palps, conceal the aperture of the mouth. They are smooth on the surface, and grooved on the opposite side close to the mouth aperture. The mouth is a large, slit-like depression placed transversely between the anterior levator muscles of the foot (Fig. 3). The mouth leads into a straight, dorso-ventrally compressed and ciliated oesophagus. The folds and depressions diversify the walls and floors of the stomach and break them into definite areas. The tissues consist largely of greenish-brown masses often termed as liver or digestive diverticula (Fig. 3). A peculiar (gelatinous) rod, the crystalline style, flattened and oblique, occupies a subcentral position anterior to where the postero-ventral fold disappears midway along the floor. The head of the crystalline style projects out of the sac where it is formed and across the cavity of the stomach where it bears against an irregular area of cuticle known as the gastric shield (Fig. 3).
The intestine may be divided into three sections, namely, the descending and ascending portions and the rectum (Fig. 3). The valvular folding of the intestinal ridge gives way to the ascending portion and curves backwards along the base of the visceral mass to the left of the descending intestine. From the point of intersection, the ascending intestine turns sharply upwards, running parallel and closely adjacent to the upper part of the descending portion (Fig. 3). The rectum runs posteriorly through the upper part of the pericardium (Fig. 3). Beyond this it curves vertically and passes around the posterior part of the adductor muscle in the median line and ends by the anus in an erectile ear-like process. The anal papilla is comparatively large and slightly curved.
The gills consist of four crescent-shaped plates, two halfgills on each side which hang down from the roof of the mantle cavity like book leaves. They represent a series of ciliated sieves, providing an efficient feeding surface. Two rows of long delicate branchial filaments are inserted at right angles along the whole length of the axis or vascular base which extends from the ventral border of the palps anteriorly curving round ventrally and posteriorly to a point opposite the anus. Its convexity extends first forwards and then downwards. Where they terminate, the mantle lobes of the two sides are slightly united by the inner mantle folds thus dividing the mantle cavity into a large inhalant chamber containing the gills and a much smaller exhalant chamber. Water enters by one and leaves by the other.
The common base of each ctenidium is a vascular ridge reaching from the anterior end of the gills. Hollow outgrowths, the inter-lamellar junctions, containing branches from the afferent vessels, convey blood from the axial trunk to the base of reflected lamellae. The blood enters the individual filaments, flows outwards to the free margin, passing over to the direct filaments, returning inward to the branchial or ctenidial axis, where it joins the efferent vessel by openings along each side. The filaments are joined chiefly by the inter-locking stiff cilia of the large ciliated discs which occur at intervals throughout their length. The normal function of the ordinary cilia on the branchiae is to create a current of water which enters the pallial chamber and passes over and through the branchial lamellae. They serve to purify the blood flowing in the filaments and to convey the food particles to the mouth.
This system consists of a heart and a series of arteries which lie above the adductor, and contained in a pericardium. The heart consists of a single ventricle and a pair of contractile thin walled auricles, one on each side. They receive blood from the body by way of the gills and mantle, and pass it to the ventricle. Back-flow of blood is prevented by valves. Blood is driven by contractions into the anterior and posterior aorta. The latter is short and supplies blood to the adductor muscle, rectum and anus. Blood is supplied to the rest of the body by the anterior aorta through a series of minor arteries. These open into the sinuses or blood spaces in which blood circulates slowly. The aorta communicates with a pair of large blood vessels that run around the margin of each mantle lobe. The deoxygenated blood is collected in veins which carry it either into the gills or excretory organs. From the kidney it is pumped into the marginal vessel of the mantle by a pair of accessory hearts. The blood from the mantle, together with that from the gills, returns to the heart through efferent branchial vein by way of auricles. The blood of the pearl oyster is colourless.
The excretory system consists of a pair of nephridia and numerous small pericardial glands projecting from the walls of the auricles. The nephridia are two large symmetrical pouch-like sacs located on either side in the hinder half of the viscero-pedal mass. Each nephridium opens into the pericardium by a wide duct and to the exterior by a minute pore. The nephridia inter-communicate by a wide channel beneath the auricles. The external renal aperture is a minute oval opening with a sphincter muscle. It opens immediately below the genital aperture within an inconspicuous lipped slit at the junction of the inner plate of the inner gill with the visceral mass at a point about midway between the ventral border of the latter and the base of the foot.
The passage connecting the right and left nephridia is wide and lies beneath the auricles. It has thin membranous walls and is bounded behind by the lower part of the pericardium and in front below with the body wall and forms part of the root of the adductor embayment of the suprabranchial chamber. The reno-pericardial tubules are a pair of wide lateral prolongations of the pre-cardiac part of the pericardium. They are thin walled, membranous and directed forwards. The aperture is a curved slit, with the concavity ventrally. The accessory pericardial glands on the walls of auricles have excretory function.
The nervous system is laterally symmetrical and has three pairs of ganglia, (1) the cerebral ganglia at the sides of the oesophagus, (2) the pedals joined to form a single ganglion at the base of the foot and (3) a pair of large visceral or parieto-splanchnic ganglia lying upon the anterior surface of the adductor. The stout paired cerebro-visceral connectives link the cerebral ganglia with the parieto-splanchnic ganglia, while a pair of cerebro pedal connectives join the cerebral ganglia with the pedal nerve mass. The cerebral ganglia are supra-oesophageal in position and a nerve cord or commissure forms the two parieto-splanchnic ganglia (Visceral ganglia). The cerebro-pedal connectives arise from the posterior and outer sides of the cerebral ganglia and run downwards within the visceral mass just behind the levator muscles of the foot to the pedal ganglion. Three principal nerves arise from the pedal ganglion and innervate the foot and the byssal gland. Each of the visceral ganglia receives from above the stout cerebro-visceral connective, the two ganglia themselves being united by a single transverse visceral commissure. Each branchial nerve leaves the ganglion at the anterior lateral corner, turns down into the base of the gills and then backwards to the posterior tips following the afferent vessels. The posterior pallial nerves emerge from the posterior end of the visceral ganglion. From the base of each, a stout nerve passes straight back and reaches the pigmented pallial sense organs of its respective side, a little anterior to the anus. The ramification of the pallial nerves in the muscular marginal region of the mantle and their anastomosing forms a complex network of nerves, the “pallial plexus.”
The sexes are separate except in occasional cases. The gonads are paired but asymmetrical. They form a thick envelop covering the stomach, liver and the stomach, and the first two sections of the intestine, connecting a greater part of the outside of the proximal portion of the viscero-pedal mass (Fig. 3). No portion of the reproductive glands extends into the foot or into the mantle. The male and female gonads are indistinguishable from external appearance in the initial stages. Both are creamy yellow in colour. In the mature stage, the male gonad is pale creamy and the female gonad yellowish creamy. The gonads of the two sexes consist of branched tubules with myriads of succate caecae, the alveoli. The spermatozoa and ova develop in these. The accumulated ripe gametes fill these alveoli and tubules and later pass into three trunks which converge into one which leads to the external genital aperture. The latter is situated dorsal to the renal aperture of the same side.
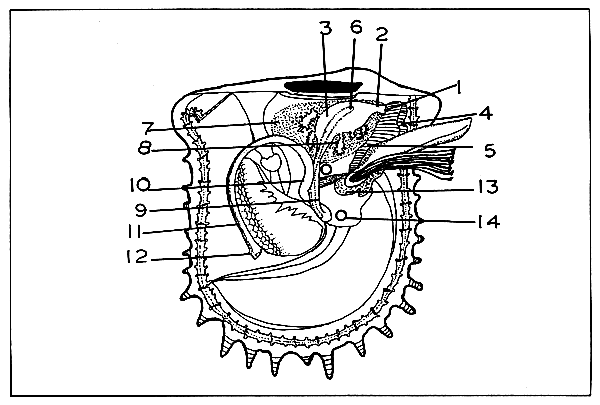
FIGURE 3. Anatomy of Pinctada fucata. 1) Mouth; 2) oesophagus; 3) stomach; 4) left labial palp; 5) left inner labial palp; 6) crystalline style; 7) liver; 8) digestive diverticula; 9) descending intestine; 10) ascending intestine; 11) rectum; 12) anal papilla; 13) byssal gland; 14) nucleus implanted in the gonad.
![]()
![]()
![]()